26A-1.イネ栽培の起源地は珠江中流域 [26A.イネ栽培の起源地は何処か]
2012年10月5日、国立遺伝研究所は、長年にわたって論争が繰り広げられてきたイネ栽培化の起源地および起源の系統について、イネのゲノム(遺伝情報)の解読を行い、その初発の起源地と栽培化のプロセスを明らかにし、論争に終止符を打つことが出来たと発表した。この研究論文は国立遺伝研究所と中国科学院との共同研究でおこなったものであり、10月3日の英科学誌『Nature』オンライン版に掲載されている。
研究チームは、世界各地から収集した栽培イネ(Oryza sativa)1083品種、その起源種とされる野生イネ(O.rufipogon)446系統を対象としてゲノム解析を行い、イネの栽培化は中国南西部を流れる珠江の中流域(広西省)で始まり、野生イネ(O.rufipogon)のかぎられた集団からジャポニカ(O.sativa japonica)が生れ、そのジャポニカと東南アジア・南アジアの野生イネとの交配によりインディカ(O.sative indica)が生れたとしている。
私は2012年1月から「稲の起源はスンダランド」をブログに投稿し、アジアの栽培イネをWaxy(粘性)遺伝子で分類して、イネの栽培化の起源地について述べている。海底に沈んだ大陸、スンダランドのデルタ地帯には野生イネのルフィポゴン(多年生)が群生していた。氷河期の終わりの海面上昇による海水の浸入のストレスで、種子をつけないルフィポゴンが穂を出し、花を咲かせ、種子を実らせた。こうして人類はイネの種子、コメを多量に手に入れた。約1万年前にはルフィポゴンの種子を栽培する人々も出て来た。熱帯においては水辺で栽培するとルフィポゴンは野生に戻り実をつけないが、陸地で栽培するとストレスを受け種子を実らせることを人々は発見し、焼畑での栽培が始まった。これがイネの栽培化の起源であり、陸稲(おかぼ)・熱帯ジャポニカの誕生である。
スンダランドでイネの栽培していた人々は、スンダランドが水没するにつれ、1万~8千年前頃に熱帯ジャポニカの種子をもって海や大陸に逃避した。海洋民族の人々は舟で中国の長江下流域に達し、イネの栽培を行った。温帯の長江下流域は、熱帯のスンダランドと比較して、夏の暑さはそれほど変わらないが、春と秋の気温差は10~12度もある。その温度差にストレスを受けた熱帯ジャポニカは、水辺で栽培しても野生に戻ることが少なく種子を実らせた。長江中・下流域で水田稲作が始まり、温帯ジャポニカが誕生した。一方、メコン川を遡った人々は、タイ東北部のコラート高原で焼畑による熱帯ジャポニカを栽培していた。そこには一年生のルフィポゴン(ニヴァラ)が自生しており、熱帯ジャポニカとニヴァラの交配の繰り返しにより、5千年前の頃にインディカが誕生した。以上が私の考えたイネの栽培化のストーリーである。
イネの栽培化は中国南西部を流れる珠江の中流域(広西省)で始まり、野生イネ・ルフィポゴン(多年生)のかぎられた集団からジャポニカが生れ、そのジャポニカと東南アジア・南アジアの野生イネとの交配によりインディカが生れたとしている国立遺伝研究所と中国科学院との共同研究の結論が真実であれば、私の考えたイネ栽培化のストーリーなど絵そらごとになってしまう。この共同研究の論文を詳細に読み、イネの栽培化の起源が珠江の中流域(広西省)であることが、真実であるかどうかを検証してみる。
26A-2.イネの起源地は遺伝距離で分かる [26A.イネ栽培の起源地は何処か]
国立遺伝学研究所の倉田のり氏は、ライフサイエンス新着論文レビューに「イネの栽培化の起源がゲノムの全域における変異比較解析により判明した」と、『Nature』に掲載された「A map of rice genome variation reveals the origin of cultivated rice 」(Huang,Kurata,at al)の解説を記載している。解説においては、野生イネのルフィポゴンについて「O.rufipogon の遺伝的な集団構造の解析を行った。系統樹解析と主成分分析により、O.rufipogonは3つのサブグープOr-I,Or-II,Or-III に分類された。・・・中国南方から採取した系統の多くはOr-IIIグループに属していた」とある。そして、栽培イネのジャポニカはルフィポゴンのOr-IIIグループ(多年性)を起源とし、インディカはルフィポゴンのOr-Iグループ(一年性)に起源をもつとしている。これらの結論は、アプローチの仕方は異なるが、既に発表されているジャポニカ・インディカの起源についての論文とほぼ同じである。
論文は「人為的な選抜による爪痕は遺伝子変異の減少と栽培化遺伝子における遺伝子頻度のかたよりとして表われる」として、55座の栽培化遺伝子を同定し、この栽培化遺伝子に焦点をあて、栽培化に関わりがあった野生イネを探している。これらを分かり易く具体的に例えれば、現在栽培しているイネの穂に実った種子(米粒)は、脱穀するまで落ちることはないが、野生イネは穂に種子を実られてもすぐに落ち(脱粒)てしまい、収穫出来る米粒は僅かである。稲作の栽培化の過程のなかで、人々はより脱粒しない品種を選択して来た。遺伝的に言えば、sh4という遺伝子に欠損が現れると脱粒が起こり難くなるそうだ。逆言えば、野生イネの集団同士を比較したとき、sh4遺伝子に欠損が多い集団が、イネの栽培化に関わりのあった野生イネであると言える。このような栽培化に関わる遺伝子のタイプをたくさん持っている野生イネが、栽培イネの起源に関係しているという考えだ。
この論文が注目を集めたのは、「イネの栽培化は中国の珠江中流域が起源であることを、ゲノム解析で解明した」と言いきったところにある。解説では「中国南方に由来するOr-IIIのサブグレードは、O.rufipogon(野生稲)のなかでもOryza sativa(栽培イネ)にもっとも近い関係にあった。・・・この55座に関して野生イネ系統を採取地別に分類したうえで、Oryza sativaとの遺伝距離を算出した結果、栽培化は中国広西地区の珠江中流域においてはじめられたことがあきらかになった」と述べている。栽培イネと同じタイプの遺伝子を持っている割合(遺伝子頻度)が高い野生イネは、栽培イネと遺伝距離が最も小さく、栽培イネの起源種であるとのことである。
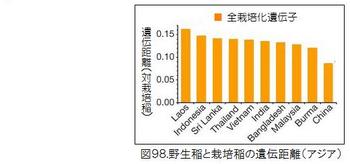
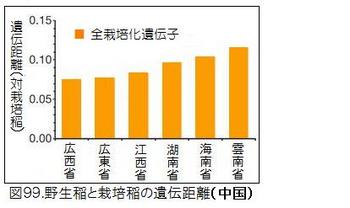
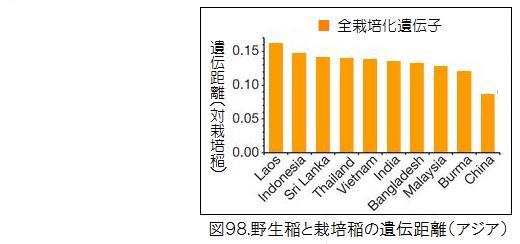
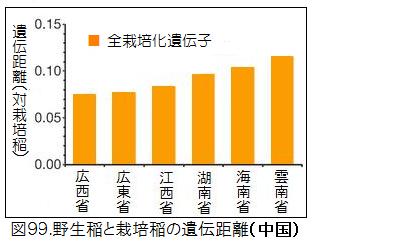
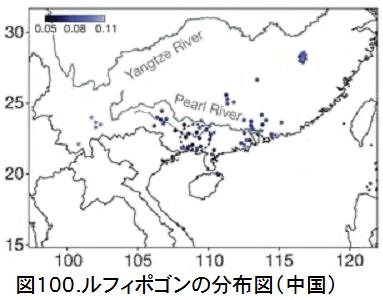
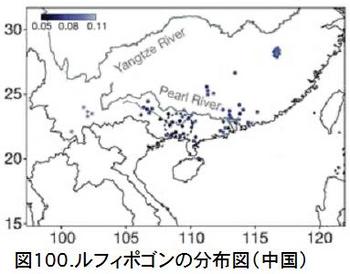 図98に示したアジアの国別の野生イネと栽培化イネの遺伝距離を見ると、中国の野生イネが際立って小さく、栽培イネに近いことが分かる。図99に示した中国内での省別の野生イネと栽培化イネの遺伝距離では、広西省・広東省が小さいことが分かる。図100は中国における野生稲ルフィポゴンの分布図である。プロットの色合いが濃いほど遺伝距離が小さく、栽培イネに遺伝的に近い。図では見えにくいが、大きな黒丸で囲っている広西省の珠江(Pearl River)中流域のプロットの色が一番濃くなっていることが分かる。ここがイネの栽培化の起源地である。
図98に示したアジアの国別の野生イネと栽培化イネの遺伝距離を見ると、中国の野生イネが際立って小さく、栽培イネに近いことが分かる。図99に示した中国内での省別の野生イネと栽培化イネの遺伝距離では、広西省・広東省が小さいことが分かる。図100は中国における野生稲ルフィポゴンの分布図である。プロットの色合いが濃いほど遺伝距離が小さく、栽培イネに遺伝的に近い。図では見えにくいが、大きな黒丸で囲っている広西省の珠江(Pearl River)中流域のプロットの色が一番濃くなっていることが分かる。ここがイネの栽培化の起源地である。
26A-3.珠江中流域の野生イネの疑問 [26A.イネ栽培の起源地は何処か]
 の『Heredity』に発表された「Genetic structure of Oryza rufipogon Griff. in China」(Wang,Zhang,at al)の論文を
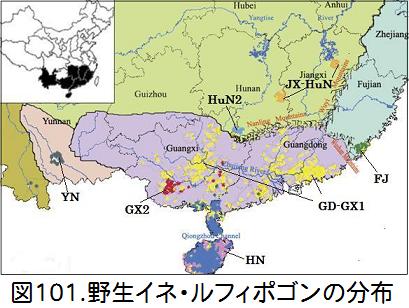
の『Heredity』に発表された「Genetic structure of Oryza rufipogon Griff. in China」(Wang,Zhang,at al)の論文を見つけた。この論文では中国の889の野生イネを対象として、その遺伝子構造を調べ、7つの地理的集団に分けている。図101にそれらを示す。黄色がGD-X1で広西・広東に分布、赤がGX2で広西に分布、ピンクがHNで海南に分布、緑がFJで福建に分布、オレンジがJX-HuNで江西・湖南に分布、青がHuN2で湖南に分布、黒がYNで雲南に分布している。
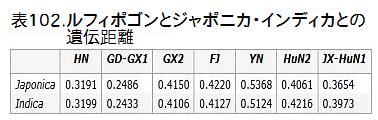
表102は、これら7つに分けられた野生イネの集団と、ジャポニカ・インディカとの遺伝子距離を示している。この表から見ると広東省・広西省に分布するGD-GX1が最も栽培イネに近い。Huang・Kurata氏の論文で栽培イネに最も近かった広西・広東の野生イネはGD-GX1のことであった。GD-GX1につい
 ては、採取された郡別に遺伝距離が算出され系統樹が作られている。この系統樹を分化の過程が分かる一般的な系統樹に書き直したのが図103である。系統樹を4等分し、それを地図上にプロットした(図104)。第一が赤●、第二が青■、第三が緑◆、第四が黄色▲である。
ては、採取された郡別に遺伝距離が算出され系統樹が作られている。この系統樹を分化の過程が分かる一般的な系統樹に書き直したのが図103である。系統樹を4等分し、それを地図上にプロットした(図104)。第一が赤●、第二が青■、第三が緑◆、第四が黄色▲である。 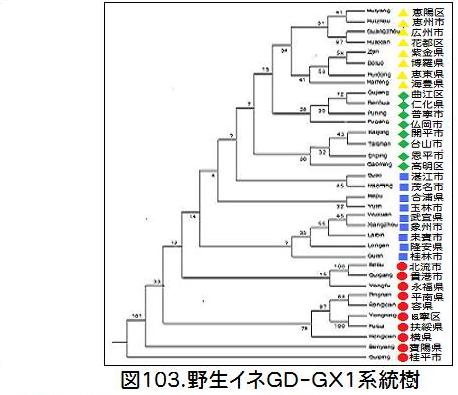

図104の大きな黒丸は、Huang・Kurata氏の論文で、ジャポニカ栽培の起源地とされた広西省の珠江中流域である。系統樹と照らし合わせると、広西の珠江中流域に出現した赤●のGD-GX1が、分化するに従いその外側に向かって青■へと拡散し、その後珠江の河口域にある支流の上流域の緑◆に広がり、最終的には珠江の河口デルタ地帯の黄色▲に定着している。拡散の情況は河下から河上へと拡散しており、Wang,Zhang氏の論文にある、水系統が関わる拡散とは思えない。花粉による拡散にしては距離が離れており、人為的な拡散のような感じがする。
私の疑問は、Wang,Zhang氏の論文のGD-GX1の系統樹で、最も分化が早いとされる珠江中流の赤●のGD-GX1が、Huang・Kurata氏の論文では、遺伝距離から見て最も栽培イネに近く、イネの栽培化の起源の野生イネと見られているという問題だ。珠江中流の赤●のGD-GX1が分化して、広東デルタ地帯の黄色▲のGD-GX1が生れたならば、広東デルタ地帯の野生イネが、栽培イネに最も近い関係にあってもよさそうに思えるのだが、事実はその逆であり、矛盾を含んでいるように思える。
26A-4.珠江中流域には銅鼓の文化があった [26A.イネ栽培の起源地は何処か]

広西省貴県(現・貴港市)に前漢時代の羅臼湾1号漢墓がある。羅臼湾との名から海岸と思いがちだが珠江中流域にあり、Huang・Kurata氏の論文でジャポニカ栽培の起源地とされた、図104で大きな黒丸をした中にある。この漢墓は出土した青銅器から、前漢武帝(179~164BC)以前であるとされている。この漢墓からは銅鼓・青銅酒器・金製品・玉璧・玉印などが出土している。図105は羅臼湾1号漢墓出土の銅鼓の写真である。
今井啓爾氏の「The Distribution of Bronze Drums of the Heger Ⅰand Pre-ⅠType」によると、広西省の珠江中流域からは、phaseⅠ(B.C2~1世紀)の羅臼湾1号漢墓をはじめとして、phaseⅢ(A.D2~?世紀)の銅鼓が非常に多
 く出土している。それらの出土地が地図にプロットされてあり、その地図と野生イネGD-GX1の採取地を重ね合わせて見た。図106を見ると、野生イネGD-GX1の採取地と銅鼓の出土地がものの見事に一致している。前漢から後漢にかけて、この地方では稲作が盛んに行われ、銅鼓を用いた豊作を願う祭祀が行われていたと想像する。それは、弥生時代の近畿から東海にかけての、銅鐸と稲作の関係と同じであるように思える。
く出土している。それらの出土地が地図にプロットされてあり、その地図と野生イネGD-GX1の採取地を重ね合わせて見た。図106を見ると、野生イネGD-GX1の採取地と銅鼓の出土地がものの見事に一致している。前漢から後漢にかけて、この地方では稲作が盛んに行われ、銅鼓を用いた豊作を願う祭祀が行われていたと想像する。それは、弥生時代の近畿から東海にかけての、銅鐸と稲作の関係と同じであるように思える。 『史記』と『漢書』には漢代の水稲栽培について、「江南は火耕水耨す」と記載されている。「耨」(ドウ)の漢字は”すきで雑草を刈り取る”という意味がある。「火耕水耨」を行ったのは江南(呉・越)と記載されているが、他の文献資料から、嶺南地方(広東・広西)やベトナムの一部の地域でも「火耕水耨」の方法で水稲栽培が行われていたことが明らかとなっている。「火耕水耨」の記載がきわめて簡単なため、具体的な内容は明確に理解出来ていないそうだが、「火耕水耨・新考」の論文を書いた彭世奨氏は、河川・湖・海岸の臨水地帯で、牛耕すのでなく火で雑草を焼き、田植えをするのでなく直播栽培で、中耕をせず水で雑草を淹死させる方法であったとしている。
 前漢から後漢にかけて、珠江中流域では水辺近くで「火耕水耨」による稲作が盛んに行われていた。その近くには野生イネGD-GX1が自生していたので、GD-GX1の花は栽培イネの花粉を他家受粉し、栽培イネの遺伝子がGD-GXに取り込まれる遺伝子流動が起こったと考える。野生イネ・ルフィポゴンの花は写真107にあるように自分の花粉で自家受粉する栽培イネと違ってオープンであり、栽培イネの花粉で他家受粉し易い構造である。これが珠江中流域の野生イネGD-GX1が最も栽培イネに近い遺伝子を多く持っていた理由であると考える。
前漢から後漢にかけて、珠江中流域では水辺近くで「火耕水耨」による稲作が盛んに行われていた。その近くには野生イネGD-GX1が自生していたので、GD-GX1の花は栽培イネの花粉を他家受粉し、栽培イネの遺伝子がGD-GXに取り込まれる遺伝子流動が起こったと考える。野生イネ・ルフィポゴンの花は写真107にあるように自分の花粉で自家受粉する栽培イネと違ってオープンであり、栽培イネの花粉で他家受粉し易い構造である。これが珠江中流域の野生イネGD-GX1が最も栽培イネに近い遺伝子を多く持っていた理由であると考える。
26A-5.珠江中流域がイネ栽培化の起源でない [26A.イネ栽培の起源地は何処か]
 Wang,Zhang 氏の論文よれば、栽培イネと野生イネ・ルフィポゴンには合計1287のhaplotype(ハプロタイプ:DNA配列)があり、その内74は栽培イネと野生イネの両者にある。その74の内11がジャポニカ特有のhaplotypeであり、38がインディカ特有のhaplotypeであるそうだ。7分類された野生イネの持つ、ジャポニカ特有のハプロタイプ(japonica haplotype)とインディカ特有のハプロタイプ(indica haplotype)の頻度(図108)が調べられている。
Wang,Zhang 氏の論文よれば、栽培イネと野生イネ・ルフィポゴンには合計1287のhaplotype(ハプロタイプ:DNA配列)があり、その内74は栽培イネと野生イネの両者にある。その74の内11がジャポニカ特有のhaplotypeであり、38がインディカ特有のhaplotypeであるそうだ。7分類された野生イネの持つ、ジャポニカ特有のハプロタイプ(japonica haplotype)とインディカ特有のハプロタイプ(indica haplotype)の頻度(図108)が調べられている。
GD-GX1においては、japonica haplotypeが約17%、indica haplotypeが約83%である。表は広東・広西に生育するGD-GX1全体の値だが、珠江中流域のものも同じ値に近いであろうと推察出来る。haplotypからすると、GD-GX1はジャポニカとインディカの両方のDNAをもっており、インディカにより近いことが分かる。人間に例えていうならば、GD-GX1にはジャポニカとインディカ両方の血が流れており、インディカの方の血が濃いということだ。
広西省貴港市の羅臼湾漢墓からは、粳(ジャポニカ)と「有客秈一石」と記載された木簡が出土している。この「客秈」とは、”外地引進的秈稲(他の地から持って来た進んだ秈稲)”と説明されている。「秈稲」とはインディカのことである。これらからして、羅臼湾漢墓がある珠江中流域は、前漢時代にジャポニカを栽培していて、そこに優良な品種のインディカ米が他の地から持ちこまれたという事であろうか。面白いことに珠江中流域からベトナムに近い地域(図106:オレンジ円)で採取されるGXは、図108にあるようにindica haplotypeが100%である。この地域からは銅鼓の出土がなく、別の文化を持つ部族が住んで、インディカを栽培していたのかも知れない。羅臼湾漢墓の木簡に書かれた、他の地区から持ち込まれた優良品種の秈(インディカ)はこの地域から来たのであろうか。前漢時代には珠江中流域で、粳(ジャポニカ)と秈(インディカ)が栽培されていたことが分かる。
前漢・後漢時代の珠江中流域の稲作は、水辺に近い所で栽培されていたので、その近くに自生していた野生イネGD-GX1は、時代のずれはあるがジャポニカとインディカ両方から花粉を受け、遺伝子流動が起こったと思える。長い稲作の歴史の中では、インディカから受けた花粉の方が多かったと考えると、図108のハプロタイプの結果が納得出来る。実際、厳文明氏の「中国稲作農業の起源」には、珠江中流域の野生イネの特性は秈(インディカ)に近いとある。
Huang・Kurata氏の論文の結論は、「イネの栽培化は中国南西部を流れる珠江の中流域で始まり、野生イネのかぎられた集団からジャポニカが生れ、そのジャポニカと東南アジア・南アジアの野生イネとの交配によりインディカが生れた」としている。その「野生イネのかぎられた集団」はWang,Zhang 氏の論文の分類でGD-GX1の一部にあたる。インディカを生んだとするジャポニカの起源種・GD-GX1に、インディカの血が既に流れているのは論理に矛盾がある。珠江中流域に生育する野生イネはジャポニカ誕生の起源種でなく、同地域がイネ栽培化の起源地ではないと考える。
26A-6.珠江中流域の野生イネは雑草イネ [26A.イネ栽培の起源地は何処か]
栽培化遺伝子に焦点を合わせて遺伝距離を出した場合、野生イネと栽培イネの遺伝距離が短いということは、野生イネが栽培イネと同じ栽培化遺伝型を多く持っているということになる。もし、栽培化の遺伝子が一挙に増えるのではなく、数千年の年月をかけて徐々にふえるのであれば、野生イネと栽培イネの遺伝距離が非常に小さいということは、その野生イネは、何千年の長い稲作の歴史の中で、栽培イネと交雑を起こし汚染(遺伝子流動)されている可能性が高いという見方も出来る。
「野生イネと栽培イネの遺伝距離が一番小さい場所が起源地」ということが成り立つのは、野生イネの集団に栽培イネからの遺伝子流動が一切なかった、あるいは有ったものは淘汰されたという場合に限り成り立つと思う。これまで述べてきたように、珠江中流域の野生イネは漢代の水際で行う「火耕水耨」の稲作により、栽培イネの花粉からの遺伝子流動を受けていたと思われる。中国の珠江中流域の野生イネの場合、遺伝距離が栽培イネに最も近いからと言って、珠江中流域がイネ栽培化の起源であると言えないと思う。
近年、中国東北部・朝鮮半島・日本の直播をする水田稲作で、雑草イネ(Weedy rice)がはびこり、米の品質を下げることが問題になってきている。この雑草イネは野生イネに栽培イネの花粉が他家受粉して出来たハイブリッド種で、遺伝的には遺伝子流動(Gene flow)が起こったと言われる。このハイブリット種は、野生イネが生息しない温帯地方では雑草イネとされるが、野生イネが生息する亜熱帯・熱帯地方では雑草イネか野生イネかの判別は付き難いそうだ。
 図109は佐藤洋一郎氏の「Origin and evolution of wild, weedy, and cultivated rice」にある、野生イネ・雑草イネ・栽培イネの関係図である。栽培イネのジャポニカ・インディカから野生イネ・ルフィポゴンに遺伝子流動が起こり、雑草イネが誕生する様子を表わしている。中国の珠江中流域に生息する野生イネGD-GX1は、2000年前に野生イネと栽培イネから出来た雑草イネであったのではないかと思える。
図109は佐藤洋一郎氏の「Origin and evolution of wild, weedy, and cultivated rice」にある、野生イネ・雑草イネ・栽培イネの関係図である。栽培イネのジャポニカ・インディカから野生イネ・ルフィポゴンに遺伝子流動が起こり、雑草イネが誕生する様子を表わしている。中国の珠江中流域に生息する野生イネGD-GX1は、2000年前に野生イネと栽培イネから出来た雑草イネであったのではないかと思える。
26A-7.中国の野生イネは海南島から始まった [26A.イネ栽培の起源地は何処か]
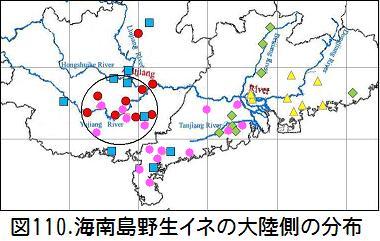
図110のピンク●は、海南島で採取される野生イネ・HNの大陸側の分布である。ピンク●のHNが海南島から大陸に渡り、赤●のGD-GX1が誕生した様子が分かる。Wang,Zhang 氏の論文の野生イネが海南から始まって広西と広東に拡散したことが明らかである。
野生イネ・ルフィポゴンは熱帯が原産地であり、1万年前の亜熱帯・温帯の中国には生育していなかったのではないかと考える。スンダランドが水没するにつれ、1万~8千年前頃に人々は海や大陸に熱帯ジャポニカの種子をもって逃避した。そのとき舟で逃避した人々には海南島に辿り着き住みついた。海南島は熱帯と亜熱帯の狭間にあり、水辺に熱帯ジャポニカの種子を植えると野生イネ・ルフィポゴンに戻って実を付けず、人々は熱帯ジャポニカの種子を焼畑で栽培するしかなかった。 その後イネ栽培は亜熱帯の大陸側に拡散した。特に北回帰線付近に位置する珠江中流域では水辺での栽培が可能となったが、野生イネに戻ることが多々あり収穫は多くなかった。
人々は気温が低いと野生イネに戻ることが少なく収穫が上がることを知り、珠江の支流の上流へと稲作が広がった。そして、南嶺山脈を越え、湖南・江西の地に辿り着いたものもあった。図101「野生イネ分布」にある「HuN2」や「JX-HuN」の先祖であろう。湖南省の玉セン岩遺跡や江西省の仙人洞遺跡の洞窟遺跡から見つかった最古(約1万年前)の栽培イネは、海南島から南嶺山脈を越えて伝わったもので、湖北省の彭頭山遺跡(約8千年前)の稲作遺跡に繋がって行く。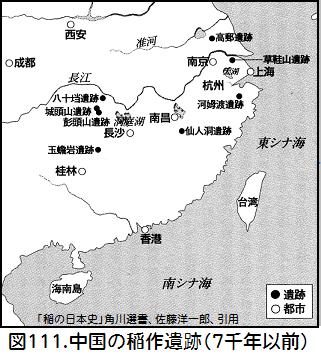 一方、海南島から舟で北上した人々は長江下流域で稲作を始めた。その遺跡が浙江省の河姆渡遺跡(7000年前)である。栽培された熱帯ジャポニカは自然環境に適応し遺伝的進化をして、水田稲作に適合する温帯ジャポニカが誕生した。河姆渡遺跡で熱帯ジャポニカと温帯ジャポニカの両方が存在するのは、温帯ジャポニカが誕生の地であるからであろう。図111は7千年以前の中国の稲作遺跡を示している。海南島から南嶺山脈を越えて拡散して行った野生イネ・ルフィポゴンが長江中流域の稲作を生み、海南島から海沿いに拡散して行った野生イネ・ルフィポゴンが長江下流域の稲作を生んだことが分かる。もちろん、これらの拡散は人間の介在によるものであった。
一方、海南島から舟で北上した人々は長江下流域で稲作を始めた。その遺跡が浙江省の河姆渡遺跡(7000年前)である。栽培された熱帯ジャポニカは自然環境に適応し遺伝的進化をして、水田稲作に適合する温帯ジャポニカが誕生した。河姆渡遺跡で熱帯ジャポニカと温帯ジャポニカの両方が存在するのは、温帯ジャポニカが誕生の地であるからであろう。図111は7千年以前の中国の稲作遺跡を示している。海南島から南嶺山脈を越えて拡散して行った野生イネ・ルフィポゴンが長江中流域の稲作を生み、海南島から海沿いに拡散して行った野生イネ・ルフィポゴンが長江下流域の稲作を生んだことが分かる。もちろん、これらの拡散は人間の介在によるものであった。
26A-8.長江下流域に野生イネはあったか [26A.イネ栽培の起源地は何処か]
 『中国の稲作起源』(1989年、渡辺武・陳文華編)にある厳文明氏の「中国稲作農業の起源」には、「栽培稲の起源、特にある特定の地域が起源の一中心地であるかどうかを探ろうとするには、その地で古い稲の遺物の発見がなくてはならない。またこれ以外にそこに野生稲が分布(あるいは過去の歴史の上で存在)してなければならず、しかもこの種の野生稲は、当地の最も古い栽培稲と遺伝学上密接な親類関係を具備していなけれ
『中国の稲作起源』(1989年、渡辺武・陳文華編)にある厳文明氏の「中国稲作農業の起源」には、「栽培稲の起源、特にある特定の地域が起源の一中心地であるかどうかを探ろうとするには、その地で古い稲の遺物の発見がなくてはならない。またこれ以外にそこに野生稲が分布(あるいは過去の歴史の上で存在)してなければならず、しかもこの種の野生稲は、当地の最も古い栽培稲と遺伝学上密接な親類関係を具備していなけれ
ばならない」としている。
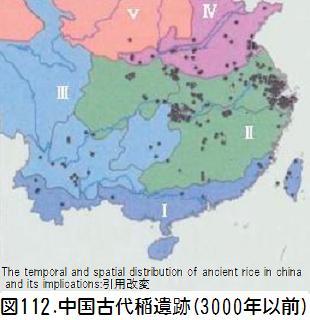
そして、「問題は、中国現存の野生稲が最も早期の栽培稲遺物の分布と重ならないことである。最古の栽培遺物は長江下流域と杭州湾地方で発見されているのに、野生稲は主に珠江流域および同緯度のその他の地域に分布している。このことは大きな矛盾である。」と述べている。図111に中国の3000年以上前の古代稲遺跡の分布図を示す。この図と現在の中国の野生イネの分布図(図101)を比較すれば、厳文明氏の話が良く分かる。
厳文明氏は、二つの可能性だけが残されているとして、一つは、華南の原始村落がまず栽培化し、その後この知識が長江下流域やその他の地方に伝えられたとする考え方。もう一つは、長江下流域に元来野生稲があり、栽培稲はまず当地で育成に成功したという考え方であるとしている。前者は考古資料の証拠が得られていない。後者については、中国の歴史文献(三国志・宋書・南史等)に「野稲」の表記があり、その分布地が最早期の栽培稲遺物が分布する地域と重なっている。文献に記録されている「野稲」は、先史時代の野生稲の遺留であるとして、後者の説を支持している。この「野稲」も気候の変化、環境の変化で滅亡の淵においやられたと記している。
佐藤洋一郎氏はイネそれもジャポニカの栽培起源地が長江中・下流域であると考えておられる。そして、浙江省の河姆渡遺跡(7000年前)の炭化米86粒を調べ、そのほとんどが栽培稲だが、その内4粒が電子顕微鏡観察で野生稲に近いと判定され、「河姆渡遺跡の田んぼにはたぶん原始的な稲が栽培されていたのであろう。そしてあるものはより野生イネに近く、またあるものは栽培化の程度が進んでいるといった不ぞろいが見られたことだろう」と述べている。 また佐藤氏は河姆渡遺跡の2粒の炭化米が熱帯ジャポニカであることをDNA分析から見つけ、試料が少ないので明確に言えないのであろうが、「想像をたくましくすれば、長江流域に起源したジャポニカは熱帯ジャポニカだったと考えられる」と述べている。長江下流域や杭州湾地方で稲作が始まった頃には、熱帯ジャポニカが栽培され、野生イネも混在していたと推測出来る。この野生イネの子孫が中国文献にある「野稲」であろう。
スンダランドから海南島を経由して、約8千年前頃長江下流域に達した人々は、熱帯ジャポニカの種子を持っていた。温帯の長江下流域では、亜熱帯の中国南部(海南・広西・広東)と比較して、熱帯ジャポニカの種子が野生イネに戻ることは少なかったが、熱帯ジャポニカに交じって野生イネ・ルフィポゴンとして生息するものもあったと思われる。この野生イネが長江下流域で誕生した温帯ジャポニカと、長い稲作の歴史の中で交雑し、雑草イネとして自生するようになった。こう考えると、長江下流域と杭州湾地方で発見されている約七千年前の栽培イネの遺物も、中国の歴史文献(三国志・宋書・南史等)に出て来る「野稲」とその分布地も、そして佐藤氏による河姆渡遺跡の炭化米の電子顕微鏡の観察とDNA分析値も、何の矛盾も起こらない。
26A-9.イネ栽培化の起源地はスンダランド [26A.イネ栽培の起源地は何処か]
上古から南宋に至る歴代の制度の沿革を記した『文献通考』には、「大中六年(852年)海陵・高郵両県(長江下流域)の百姓、官河中より異稲を漉得して煮食す。呼びて聖米と為す」とある。この「異稲」が「野稲」で野生イネとされている。「漉」(こす)の漢字からして、「異稲」の籾は稲刈りをしたのでなく、脱粒して河に漂っていたのを掬い採ったであろう。野生イネは脱粒性が高いので、完熟まで待てば脱粒していまい、脱粒前に刈り取れば未熟である。人類が初めて野生イネの実を手にしたのは、稲刈りでなく水中に漂う籾を掬いとることであったと考える。
人々が野生イネの実・米を食糧としたのは、多量の籾が採れたからだと思う。少量の籾ならば手間をかけて籾殻を取り除き、また土器を用いて煮炊きしなければ食べられないものを、食糧にしようとは思わなかったと考える。水辺に群生している野生イネ・ルフィポゴンは、普通は種子を実らせない。その野生イネがいっせいに穂を出し、花を咲かせ、種子を実らせ、そして脱落し水中に多量に籾が漂う現象が起こり、人々は初めて野生イネの籾を多量に手に入れる事が出来たと思う。その現象が氷河期の終わりの1万5千年から1万年前、海面の上昇で沈みゆく広大なスンダランド大陸に起こったと考える。
 野生イネの籾を多量に手に入れた人々は、籾を脱穀・煮炊きすれば食べられること、また籾を乾燥しておけば長期に保存出来る食
野生イネの籾を多量に手に入れた人々は、籾を脱穀・煮炊きすれば食べられること、また籾を乾燥しておけば長期に保存出来る食糧となることを知ったのであろう。スンダランドの海面は徐々に上昇
したので、野生イネの実り現象は河上に向かって数千年続いたのかも知れない。その間に野生イネの籾を陸地に播けば、穂を出し、花を咲かせ、種子を実らせること、また野山を焼いて籾を播けば収穫が上がることを知ったと思う。これが焼畑による稲栽培化の起源であり、陸稲(おかぼ)・熱帯ジャポニカの誕生である。ルフィポゴンの子孫である熱帯ジャポニカが、多年草であるにも関わらず陸稲であるのはこのためであると考える。 スンダランド(図50)がイネ栽培化の起源地であるとの考えは変わっていない。



