25-1.葉緑体DNAは母系の遺伝子 [25.インディカの起源を探る]
 植物の細胞では、核・ミトコンドリアと葉緑体にDNAがある。ゲノム(全部のDNA)を構成する塩基対の数は、イネの核ゲノムでは約4億対、ミトコンドリアゲノムは約5千万対、葉緑体ゲノムは約14万対である。そのため、葉緑体のDNAの完全解読が、核やミトコンドリアに先駆けて1988年になされている。核のDNAは父親と母親の双方の遺伝子を受け継ぐが、葉緑体のDNAは母親のみの遺伝子を受け継いでいる。そのため、葉緑体のDNAを分析すれば、母方のルーツを探ることが出来る。
植物の細胞では、核・ミトコンドリアと葉緑体にDNAがある。ゲノム(全部のDNA)を構成する塩基対の数は、イネの核ゲノムでは約4億対、ミトコンドリアゲノムは約5千万対、葉緑体ゲノムは約14万対である。そのため、葉緑体のDNAの完全解読が、核やミトコンドリアに先駆けて1988年になされている。核のDNAは父親と母親の双方の遺伝子を受け継ぐが、葉緑体のDNAは母親のみの遺伝子を受け継いでいる。そのため、葉緑体のDNAを分析すれば、母方のルーツを探ることが出来る。1998年に千葉大学の中村郁郎氏等は、栽培イネのジャポニカとインディカ、そして、栽培イネに最も近い野生イネの多年生ルフィポゴンと1年生ルフィポゴンの、葉緑体のDNAを分析している。 分析された項目は、葉緑体のORF100領域での塩基の欠
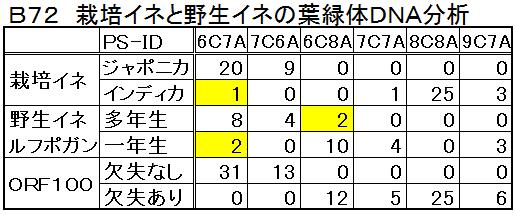 失とPS-ID領域の塩基の並びとである。それらの結果を表B72に示す。表のルフィポゴンにある黄色マークの3種を別にすると、ジャポニカと多年生ルフィポゴンには、ORF100領域の欠失はなく、インディカと1年生ルフィポゴンには、ORF100の欠失があるといえる。
失とPS-ID領域の塩基の並びとである。それらの結果を表B72に示す。表のルフィポゴンにある黄色マークの3種を別にすると、ジャポニカと多年生ルフィポゴンには、ORF100領域の欠失はなく、インディカと1年生ルフィポゴンには、ORF100の欠失があるといえる。 なお、黄色マークの「欠失のないインディカ」、「欠失のない1年生ルフィポゴン」、「決失のある多年生ルフポゴン」とは、数代に渡って他家受粉(他品種の花粉で受粉)したために起こる、核置換型といわれる、葉緑体が母系で核が父系の雑種である。イネは花の構造や開花時期の仕方が、自家受粉するように出来ていて、葉緑体と核の遺伝子は、世代が変わっても同じである。しかし自然界において1%程度、他家受粉する場合があるそうだ。
アジアのイネ属は栽培イネの2種と野生イネの7種である。その中でゲノムタイプが「AA」であるものは、栽培イネのジャポニカとインディカ、そして野生イネのルフィポゴンのみである。一方、イネ属の中で「ORF100の欠失」のあるものは、インディカと1年生ルフィポゴンのみである。これらからすると、ジャポニカの先祖は多年生ルフィポゴン、インディカの先祖は一年生ルフィポゴンと考えるのが自然である。なお、一年生ルフィポゴンはニヴァラと称されており、以後、一年生ルフィポゴンをニヴァラ、多年生ルフィポゴンをルフィポゴンと呼ぶ。
PS-ID領域のジャポニカとルフィポゴンの細胞質型式は、「7C6A ・6C7A」の2型式。別の研究によると熱帯ジャポニカは「7C6A ・6C7A」で、温帯ジャポニカは「6C7A」のみである。
7C6A;TAACCCCCCCAAAAAAGTA :ジャポニカ・ルフィポゴン
6C7A;TAACCCCCCAAAAAAAGTA :ジャポニカ・ルフィポゴン
インディカと一年生ルフィポゴンの細胞質型式は、「9C7A ・8C8A・7C7A・6C8A」の4型式ある。
9C7A;TAACCCCCCCCCAAAAAAAGTA:インディカ・ニヴァラ
8C8A:TAACCCCCCCCAAAAAAAAGTA :インディカ
7C7A:TAACCCCCCCAAAAAAAGTA :インディカ・ニヴァラ
6C8A:TAACCCCCCCCAAAAAAGTA :ニヴァラ
25-2.葉緑体DNAも進化している [25.インディカの起源を探る]
 葉緑体のDNAは、母親のみの遺伝子を受け継いでいるので、遺伝子は変わらないと考えられていた。2004年にミトコンドリアゲノムが解読され、その働きが解明出来るようになり、図B73に示すように、葉緑体からDNAの断片(塩基)が欠落し、ミミトコンドリアを通じて核に、あるいは葉緑体から直接核に、塩基が移動していることが分かってきた。図は「ミトコンドリアゲノム再編成に関する研究」門脇幸一(NIS)、2004年を引用した。葉緑体のDNAは、母系の先祖と違って来ることも在りうるのだ。
葉緑体のDNAは、母親のみの遺伝子を受け継いでいるので、遺伝子は変わらないと考えられていた。2004年にミトコンドリアゲノムが解読され、その働きが解明出来るようになり、図B73に示すように、葉緑体からDNAの断片(塩基)が欠落し、ミミトコンドリアを通じて核に、あるいは葉緑体から直接核に、塩基が移動していることが分かってきた。図は「ミトコンドリアゲノム再編成に関する研究」門脇幸一(NIS)、2004年を引用した。葉緑体のDNAは、母系の先祖と違って来ることも在りうるのだ。 進化が進むと多様性が進むというのが、進化の原則である。それならば、葉緑体DNAにおいても、先祖はより集約された形であったと私は考える。PS-IDにおいて、ジャポニカとルフィポゴンの先祖、インディカとニヴァラルの先祖の細胞質型は、次に示すようにそれぞれ一つの型式であったと考える。
ジャポニカとルフィポゴンの先祖の葉緑体DNAの細胞質型式は「7C7A」である。
7C7A;TAACCCCCCCAAAAAAAGTA
インディカとニヴァラの先祖の細胞質は「9C8A」である。
9C8A;TAACCCCCCCCCAAAAAAAAGTA
これらから「C」、または「A」の塩基が欠落したと考えると、ジャポニカとルフィポゴンの「6C7A・7C6A」が生れ、インディカの「9C7A・8C8A・7C7A」も、ニヴァラの「9C7A・7C7A・6C8A」も生れて来る。
2008年にタイのMahasarakham大学が、タイの栽培イネの在来種74種について、PS-ID領域における細胞質型の分析を行っている。この74種の中に、ORF100領域の欠失がない、正常のジャポニカに「7C7A」が4品種、ORF100領域に欠失のある、正常のインディカに「9C8A」が1品種あるのを見つけた。これらより、栽培イネの先祖が野生イネならば、その先祖の葉緑体DNAの細胞質型は、「7C7A」のルフィポゴンと、「9C8A」のニヴァラが存在していたと私は推察する。
25-3.栽培イネと野生イネのルーツ [25.インディカの起源を探る]
 東京大学の大坪栄一氏等は、イネのWaxy遺伝子内に散在している動き回る遺伝子、レトロポゾンp-SENE1を分析し、イネの進化について調べている。動き回る遺伝子について最初に唱えた、マクリントックという女性科学者は、ノーベル賞を受賞したくらいであるから、これを理解することはさておいて、その研究の成果としての栽培イネと野生イネの系統樹を図B74に掲げる。この図は「Polyphyletic Origin of Cultivated Rice(栽培イネの多系統起源))」2003年のCheng等の論文から引用した。
東京大学の大坪栄一氏等は、イネのWaxy遺伝子内に散在している動き回る遺伝子、レトロポゾンp-SENE1を分析し、イネの進化について調べている。動き回る遺伝子について最初に唱えた、マクリントックという女性科学者は、ノーベル賞を受賞したくらいであるから、これを理解することはさておいて、その研究の成果としての栽培イネと野生イネの系統樹を図B74に掲げる。この図は「Polyphyletic Origin of Cultivated Rice(栽培イネの多系統起源))」2003年のCheng等の論文から引用した。 図は分かり易くするため品種名は国名に変え、中間的な品種は省いている。また、多年生のルフィポゴンはルフィポゴン、一年生ルフィポゴンはニヴァラと書き変えている。栽培イネのジャポニカ(青)とインディカ(紫)、野生イネのルフィポゴン(赤)とニヴァラ(緑)の系統樹を見て分かることは、ジャポニカは野生イネのルフィポゴンと先祖が同じであり、インディカはニヴァラと先祖が同じであるということだ。 面白い事に、見方によっては一部のインディカはニヴァラより先祖に近くなっている。
野生イネも1万年の間に、突然変異により多様化している。ジャポニカの先祖は、現在のルフィポゴンではなく「7C7A」のルフィポゴンで、インディカの先祖は、現在のニヴァラではなく「9C8A」のニヴァラであったと考える。なお、「紫」のインディカの枝に「青」のジャポニカが4個あり、「青」のジャポニカの枝に「紫」のインディカが1個ある。そして、「赤」のルフィポゴンの枝に「緑」のニヴァラが1個ある。これは、前節25-2で述べた「置換型」で、葉緑体DNAと核DNAとが異なる品種であるためだと推察した。この「置換型」がインディカの枝に多いということが、インディカの出生の秘密を解くカギとなった。
25-4.インディカの起源はタイ東北部 [25.インディカの起源を探る]
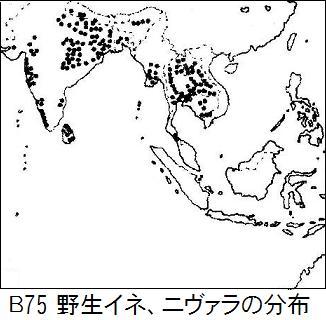 ニヴァラは一年生で、雨季と乾季がはっきりしている所に生息し、乾季には。花を咲かせ種子を実らせる。種子は脱落し、次の雨季が来るまで休眠し発芽しない。すぐ発芽すると、水がないから枯れてしまい繁殖できないからだ。図B75に見られるように、ニヴァラはインドシナ半島とインド大陸の熱帯サバンナ気候に多く生息している。 前節の図B74の系統樹において、緑色のニヴァラではインドのニヴァラは枝の先にあり、タイのニヴァラはそれよりも幹に近い所にある。このことより、インディカの先祖は、インドではなく
ニヴァラは一年生で、雨季と乾季がはっきりしている所に生息し、乾季には。花を咲かせ種子を実らせる。種子は脱落し、次の雨季が来るまで休眠し発芽しない。すぐ発芽すると、水がないから枯れてしまい繁殖できないからだ。図B75に見られるように、ニヴァラはインドシナ半島とインド大陸の熱帯サバンナ気候に多く生息している。 前節の図B74の系統樹において、緑色のニヴァラではインドのニヴァラは枝の先にあり、タイのニヴァラはそれよりも幹に近い所にある。このことより、インディカの先祖は、インドではなくタイに生息するニヴァラに近いことが分かる。
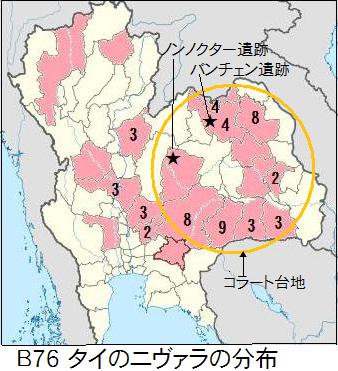 タイのKasetsart Universtyは、2009年に「香り米」の遺伝子をもたらした野生イネを同定するために、ルフィポゴン184種、ニヴァラ60種を調査している。ニヴァラの採集地を図B76に示した。数字は採集品種数、1は未記入。これらから見ると、ニヴァラはタイの東北部にあるコラート台地に多く生息している。コラート台地は標高200m程度の広大な台地である。私は南北と東西に二度横断したが、その印象は「たいらランド、タイランド」であった。台地にはメコン川の支流が流れている。
タイのKasetsart Universtyは、2009年に「香り米」の遺伝子をもたらした野生イネを同定するために、ルフィポゴン184種、ニヴァラ60種を調査している。ニヴァラの採集地を図B76に示した。数字は採集品種数、1は未記入。これらから見ると、ニヴァラはタイの東北部にあるコラート台地に多く生息している。コラート台地は標高200m程度の広大な台地である。私は南北と東西に二度横断したが、その印象は「たいらランド、タイランド」であった。台地にはメコン川の支流が流れている。 スンダランドのほとんどが水没した頃、熱帯ジャポニカのウルチ米を携えてメコン川を遡った人々は、ラオス北部やタイ北部に定住し、焼畑による稲作を始めた。その人々の中には、コラー台地の北部に定住した人々もいた。バンチェン遺跡やノンノクター遺跡がある地域である。こと辺りの川辺は、雨季には浸水するが乾季には乾燥して、人々が栽培した陸稲の熱帯ジャポニカのウルチ米は、野生のルフポゴンに戻ることなく実った。

熱帯ジャポニカを植えた土地には、野生イネのニヴァラが生えていたため、ニヴァラの花は熱帯ジャポニカの花粉で受粉する他家受粉の機会が多くなった。数代に渡って他家受粉を繰り返して行くと、葉緑体の遺伝子は母系のニヴァラであるが、核の遺伝子は父系の熱帯ジャポニカのウルチである核置換型品種、インディカが誕生した。図B77に核置換型品種の出来る過程を示す。この図は「稲作の起源」池橋宏より引用し、赤字は加筆した。
自然界ではニヴァラとルフポゴンも棲み分けが出来ていて、交雑することがなかったのが、栽培ということで人間が介在し、棲み分けを無くしたために、新しい品種のインディカが誕生したのである。インディカはたわわに実り、脱粒が少なく多くの米が収穫出来るので、人々はインディカを選別して栽培するようになった。インディカのウルチの誕生は、熱帯ジャポニカのモチと同じ5000年前の頃と考える。
25-5.インディカ米にもモチ米がある [25.インディカの起源を探る]
 私はタイに駐在したことがあるが、タイには酒飲みでも喜んで食べる「カオニャオマムアン」というスイ―ツがある。「カオニャオ」はモチ米で、「マムアン」がマンゴであり、モチ米のおこわにマンゴを添え、塩味のココナツミルクをかけてある。聞いただけではチョット遠慮したいが、食べてみるとマイルドで美味しい。このモチ米はインディカ米である。 モチ米は温帯ジャポニカだけなく、熱帯ジャポニカもインディカにもある。モチ米に対応するのがウルチ米。三品種それぞれにウルチ米がある。日本人が日常食べているのが、温帯ジャポニカのウルチ米である。
私はタイに駐在したことがあるが、タイには酒飲みでも喜んで食べる「カオニャオマムアン」というスイ―ツがある。「カオニャオ」はモチ米で、「マムアン」がマンゴであり、モチ米のおこわにマンゴを添え、塩味のココナツミルクをかけてある。聞いただけではチョット遠慮したいが、食べてみるとマイルドで美味しい。このモチ米はインディカ米である。 モチ米は温帯ジャポニカだけなく、熱帯ジャポニカもインディカにもある。モチ米に対応するのがウルチ米。三品種それぞれにウルチ米がある。日本人が日常食べているのが、温帯ジャポニカのウルチ米である。 モチ米のデンプン成分はアミロペクチンだけだが、ウルチ米には20パーセント程度アミロースという成分が入っている。アミロースの割合が増えるほど米の粘り気がなくなってくる。それでは、アミロースの割合が同じであるジャポニカとインディカでは、同じ粘り気になるかと言えば、インディカの方がパサパサしているそうだ。その原因はジャポニカとインディカのアミロクプチンの構造の違いによるものだそうだ。
25-6.インディカは天然ハイブリッド米 [25.インディカの起源を探る]
インディカが生れたバンチェン遺跡やノンノクター遺跡の地域には、熱帯ジャポニカのモチも栽培されていたので、インディカのウルチ、熱帯ジャポニカのモチ、そして野生イネのニヴァラの交配が起こり、多くのインディカ品種が誕生した。
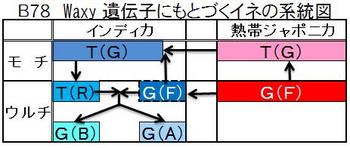 図B78はWaxy遺伝子から見たイネの系統図である。一年生の野生イネのニヴァラの花に、熱帯ジャポニカのウルチG(F)の花粉が数代に渡り受粉して、インディカのウルチG(F)が生れ、その花に熱帯ジャポニカのモチT(G)の花粉が受粉して、インディカのモチT(G)が生れている。そして、その花にニヴァラの花粉が受粉して、インディカのT(R)が生れている。T(R)はウルチとモチの中間的な形質である。
図B78はWaxy遺伝子から見たイネの系統図である。一年生の野生イネのニヴァラの花に、熱帯ジャポニカのウルチG(F)の花粉が数代に渡り受粉して、インディカのウルチG(F)が生れ、その花に熱帯ジャポニカのモチT(G)の花粉が受粉して、インディカのモチT(G)が生れている。そして、その花にニヴァラの花粉が受粉して、インディカのT(R)が生れている。T(R)はウルチとモチの中間的な形質である。
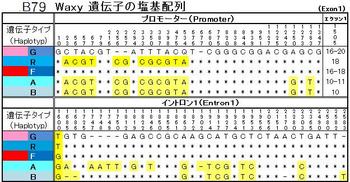 図B79Waxy遺伝子の塩基配列で見られるように、「G」と「F」においては、熱帯ジャポニカとインディカは全く同じで、イントロン最初の塩基がただひとつG→Tに変わり、ウルチがモチに変っている。インディカの「R」とモチ「G」とは、イントロンの塩基25個は全く同じで、プロモーターの塩基32個の内12個が違っている。この大きな違いは、野生イネのニヴァラの遺伝子が、形質を決める核の遺伝子と混ざったためであると推察した。
図B79Waxy遺伝子の塩基配列で見られるように、「G」と「F」においては、熱帯ジャポニカとインディカは全く同じで、イントロン最初の塩基がただひとつG→Tに変わり、ウルチがモチに変っている。インディカの「R」とモチ「G」とは、イントロンの塩基25個は全く同じで、プロモーターの塩基32個の内12個が違っている。この大きな違いは、野生イネのニヴァラの遺伝子が、形質を決める核の遺伝子と混ざったためであると推察した。
インディカのT(R)はインディカのウルチG(F)と交配し、インディカのウルチの主力品種G(A)・G(B)が生れている。野生イネのニヴァラの遺伝子は、この主力品種G(A)・G(B)に受け継がれることになる。現在、ハイブリッド米と言って、縁遠い品種を交配させ雑種の強さを生かす品種改良の研究がある。その中には栽培イネと野生イネの交配も行われている。インディカは自然が作ったハイブリッド米である。インディカがジャポニカに比べ、多くの地域で栽培されているのは、野生の血が入った生命力のある、そして収穫の多い品種だったからであろう。
 Waxy遺伝子に基づいたインディカの系統図に、その栽培地を記し、図B80に示した。インディカのウルチG(F)にも、モチT(G)にもタイの名がある。インディカのT(R)には、バングラデッシュとインドネシアだけでタイの名がないが、きっとタイの在来種にもT(R)があると考える。インディカの主力品種のG(A)は中国南部とインドシナ半島で栽培され、G(B)はインド・スリランカなどインド大陸で栽培されている。タイの東北部のコラート台地で誕生したと比定したインディカが、どのようなルートでアジアに広がったかを考えてみる。
Waxy遺伝子に基づいたインディカの系統図に、その栽培地を記し、図B80に示した。インディカのウルチG(F)にも、モチT(G)にもタイの名がある。インディカのT(R)には、バングラデッシュとインドネシアだけでタイの名がないが、きっとタイの在来種にもT(R)があると考える。インディカの主力品種のG(A)は中国南部とインドシナ半島で栽培され、G(B)はインド・スリランカなどインド大陸で栽培されている。タイの東北部のコラート台地で誕生したと比定したインディカが、どのようなルートでアジアに広がったかを考えてみる。
 図B78はWaxy遺伝子から見たイネの系統図である。一年生の野生イネのニヴァラの花に、熱帯ジャポニカのウルチG(F)の花粉が数代に渡り受粉して、インディカのウルチG(F)が生れ、その花に熱帯ジャポニカのモチT(G)の花粉が受粉して、インディカのモチT(G)が生れている。そして、その花にニヴァラの花粉が受粉して、インディカのT(R)が生れている。T(R)はウルチとモチの中間的な形質である。
図B78はWaxy遺伝子から見たイネの系統図である。一年生の野生イネのニヴァラの花に、熱帯ジャポニカのウルチG(F)の花粉が数代に渡り受粉して、インディカのウルチG(F)が生れ、その花に熱帯ジャポニカのモチT(G)の花粉が受粉して、インディカのモチT(G)が生れている。そして、その花にニヴァラの花粉が受粉して、インディカのT(R)が生れている。T(R)はウルチとモチの中間的な形質である。  図B79Waxy遺伝子の塩基配列で見られるように、「G」と「F」においては、熱帯ジャポニカとインディカは全く同じで、イントロン最初の塩基がただひとつG→Tに変わり、ウルチがモチに変っている。インディカの「R」とモチ「G」とは、イントロンの塩基25個は全く同じで、プロモーターの塩基32個の内12個が違っている。この大きな違いは、野生イネのニヴァラの遺伝子が、形質を決める核の遺伝子と混ざったためであると推察した。
図B79Waxy遺伝子の塩基配列で見られるように、「G」と「F」においては、熱帯ジャポニカとインディカは全く同じで、イントロン最初の塩基がただひとつG→Tに変わり、ウルチがモチに変っている。インディカの「R」とモチ「G」とは、イントロンの塩基25個は全く同じで、プロモーターの塩基32個の内12個が違っている。この大きな違いは、野生イネのニヴァラの遺伝子が、形質を決める核の遺伝子と混ざったためであると推察した。 インディカのT(R)はインディカのウルチG(F)と交配し、インディカのウルチの主力品種G(A)・G(B)が生れている。野生イネのニヴァラの遺伝子は、この主力品種G(A)・G(B)に受け継がれることになる。現在、ハイブリッド米と言って、縁遠い品種を交配させ雑種の強さを生かす品種改良の研究がある。その中には栽培イネと野生イネの交配も行われている。インディカは自然が作ったハイブリッド米である。インディカがジャポニカに比べ、多くの地域で栽培されているのは、野生の血が入った生命力のある、そして収穫の多い品種だったからであろう。
 Waxy遺伝子に基づいたインディカの系統図に、その栽培地を記し、図B80に示した。インディカのウルチG(F)にも、モチT(G)にもタイの名がある。インディカのT(R)には、バングラデッシュとインドネシアだけでタイの名がないが、きっとタイの在来種にもT(R)があると考える。インディカの主力品種のG(A)は中国南部とインドシナ半島で栽培され、G(B)はインド・スリランカなどインド大陸で栽培されている。タイの東北部のコラート台地で誕生したと比定したインディカが、どのようなルートでアジアに広がったかを考えてみる。
Waxy遺伝子に基づいたインディカの系統図に、その栽培地を記し、図B80に示した。インディカのウルチG(F)にも、モチT(G)にもタイの名がある。インディカのT(R)には、バングラデッシュとインドネシアだけでタイの名がないが、きっとタイの在来種にもT(R)があると考える。インディカの主力品種のG(A)は中国南部とインドシナ半島で栽培され、G(B)はインド・スリランカなどインド大陸で栽培されている。タイの東北部のコラート台地で誕生したと比定したインディカが、どのようなルートでアジアに広がったかを考えてみる。



