23-1.世界最古の稲作遺跡 [23.稲の起源はスンダランド]
1973年に中国の長江下流域、浙江省の省都杭州の東100キロにある河姆渡(かぼと)村で多量の稲作遺物が発見された。この遺跡からは150トンにも及ぶイネや、イネの栽培を裏付ける生産道具や生活道具が2838点も出土している。そして、C14炭素年代測定により、この遺跡が7000年前のものと判明した。
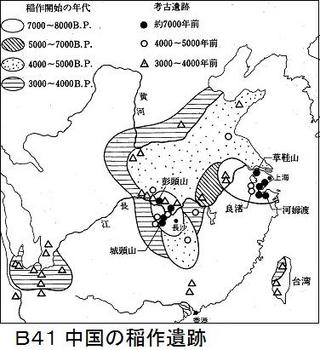 中国では河姆渡遺跡と同時代の稲作遺跡が、1980年代以降に次々と発見されている。長江下流域では杭州北方の羅家角遺跡、太湖がある江蘇省蘇州付近の草鞋山遺跡などである。また、長江中流域の洞庭湖のある湖南省では、彭頭山遺跡や城頭山遺跡である。図B41に「中国の稲作遺跡」を示した。この図は「DNAが語る稲作文明」(佐藤洋一郎)から引用した。これらの稲作遺跡の発見により、稲作の起源地は長江流域であるということが唱えられ、それまで稲作起源地とされてきた雲南地方の稲作遺跡は、いまのところ4000年前のものであることからして、「アッサム・雲南」説が覆されたのである。
中国では河姆渡遺跡と同時代の稲作遺跡が、1980年代以降に次々と発見されている。長江下流域では杭州北方の羅家角遺跡、太湖がある江蘇省蘇州付近の草鞋山遺跡などである。また、長江中流域の洞庭湖のある湖南省では、彭頭山遺跡や城頭山遺跡である。図B41に「中国の稲作遺跡」を示した。この図は「DNAが語る稲作文明」(佐藤洋一郎)から引用した。これらの稲作遺跡の発見により、稲作の起源地は長江流域であるということが唱えられ、それまで稲作起源地とされてきた雲南地方の稲作遺跡は、いまのところ4000年前のものであることからして、「アッサム・雲南」説が覆されたのである。
これら6000年前以前の稲作遺跡で作られていたイネは、何の品種であったかという事は議論が分れている。中国の学者の多くは、「稲作の起源はインディカもジャポニカも中国」という考えである。その論拠の一つは、河姆渡遺跡などから出土した米粒のかたちが色々あることで、インディカとジャポニカの両方が混在していた証拠と考えられている。
佐藤氏は中国の古代稲作遺跡(6000千年前以前)より出土した炭化米22粒のDNA分析を行い、22粒全部がジャポニカで、その内の4粒が熱帯ジャポニカ(残りは熱帯か温帯ジャポニカか不明)であったことより、稲作の起源地は中国の長江流域であるが、そこで栽培されたイネはジャポニカであったと考え、インディカは別の場所が起源地であると考えている。ただ、インディカの起源地は現在でも特定出来ていない。イネのプラントオパール分析を開発した藤原氏は、中国と共同研究を行い、草鞋山遺跡の水田遺構を検出するとともに、その栽培されたイネがジャポニカであったと判定している。
中国の古代稲作の品種が何であったかという問題は、ただその品種を特定するという問題だけではない。イネの先祖の系譜を解き明かすための、大きな問題なのだ。一昔前では、これらの問題を解き明かす手法は、育種を通じての形態的な研究が中心であったが、現在ではDNA分析が用いられている。「ジャポニカとインディカの先祖は、同じ野生イネか、それとも別々の野生イネか。稲作の起源地は中国か、それともジャポニカは中国、インディカはインドか。ジャポニカとインディカどちらが先に生れたか。温帯ジャポニカと熱帯ジャポニカではどちらが先か。」、現在の栽培イネや野生イネのDNAを調べることで、先祖の系譜を知ることが出来るのだ。ただ、今まで多くの研究者がDNA分析の結果を報告しているが、まだ明解な答えは出ていない。
 中国では河姆渡遺跡と同時代の稲作遺跡が、1980年代以降に次々と発見されている。長江下流域では杭州北方の羅家角遺跡、太湖がある江蘇省蘇州付近の草鞋山遺跡などである。また、長江中流域の洞庭湖のある湖南省では、彭頭山遺跡や城頭山遺跡である。図B41に「中国の稲作遺跡」を示した。この図は「DNAが語る稲作文明」(佐藤洋一郎)から引用した。これらの稲作遺跡の発見により、稲作の起源地は長江流域であるということが唱えられ、それまで稲作起源地とされてきた雲南地方の稲作遺跡は、いまのところ4000年前のものであることからして、「アッサム・雲南」説が覆されたのである。
中国では河姆渡遺跡と同時代の稲作遺跡が、1980年代以降に次々と発見されている。長江下流域では杭州北方の羅家角遺跡、太湖がある江蘇省蘇州付近の草鞋山遺跡などである。また、長江中流域の洞庭湖のある湖南省では、彭頭山遺跡や城頭山遺跡である。図B41に「中国の稲作遺跡」を示した。この図は「DNAが語る稲作文明」(佐藤洋一郎)から引用した。これらの稲作遺跡の発見により、稲作の起源地は長江流域であるということが唱えられ、それまで稲作起源地とされてきた雲南地方の稲作遺跡は、いまのところ4000年前のものであることからして、「アッサム・雲南」説が覆されたのである。 これら6000年前以前の稲作遺跡で作られていたイネは、何の品種であったかという事は議論が分れている。中国の学者の多くは、「稲作の起源はインディカもジャポニカも中国」という考えである。その論拠の一つは、河姆渡遺跡などから出土した米粒のかたちが色々あることで、インディカとジャポニカの両方が混在していた証拠と考えられている。
佐藤氏は中国の古代稲作遺跡(6000千年前以前)より出土した炭化米22粒のDNA分析を行い、22粒全部がジャポニカで、その内の4粒が熱帯ジャポニカ(残りは熱帯か温帯ジャポニカか不明)であったことより、稲作の起源地は中国の長江流域であるが、そこで栽培されたイネはジャポニカであったと考え、インディカは別の場所が起源地であると考えている。ただ、インディカの起源地は現在でも特定出来ていない。イネのプラントオパール分析を開発した藤原氏は、中国と共同研究を行い、草鞋山遺跡の水田遺構を検出するとともに、その栽培されたイネがジャポニカであったと判定している。
中国の古代稲作の品種が何であったかという問題は、ただその品種を特定するという問題だけではない。イネの先祖の系譜を解き明かすための、大きな問題なのだ。一昔前では、これらの問題を解き明かす手法は、育種を通じての形態的な研究が中心であったが、現在ではDNA分析が用いられている。「ジャポニカとインディカの先祖は、同じ野生イネか、それとも別々の野生イネか。稲作の起源地は中国か、それともジャポニカは中国、インディカはインドか。ジャポニカとインディカどちらが先に生れたか。温帯ジャポニカと熱帯ジャポニカではどちらが先か。」、現在の栽培イネや野生イネのDNAを調べることで、先祖の系譜を知ることが出来るのだ。ただ、今まで多くの研究者がDNA分析の結果を報告しているが、まだ明解な答えは出ていない。
23-2.イネの起源は熱帯ジャポニカ [23.稲の起源はスンダランド]
2002年12月に小泉総理は世界に向けて、「イネのゲノム(すべてのDNA)の解読が終了した」と宣言した。解読が終了したということは、DNAに書きこまれている記号(塩基配列)を全て読んだということで、その塩基配列がどんな働きをしているのか、全て解ったということではない。いま世界の多くの学者が、イネのDNAのどの部分の遺伝子の塩基配列が、何の働きをしているのか、研究の鎬(しのぎ)を削っている。
イネのDNA研究の先頭を走っているのが、農業生物資源研究所(NIAS)である。そのNIASが2008年7月、コメの大きさを決める遺伝子(qSW5)を発見したと発表した。それまでに見つけていたイネの脱粒性(穂からの籾のこぼれ易さ)の遺伝子(qSH1)と、炊いたコメのもちもち感を決める遺伝子(Waxy)と合わせて、三つの遺伝子の変化について、アジア15ヶ国のイネ、インディカ31種、ジャポニカ111種のDNA分析を行っている。
その結果、インディカでは、qSW5・qSH1・WaxyのDNAの変化はほとんど見られなかったが、ジャポニカでは、色々の変化のパターンが見られ、インディカとジャポニカは、別々の過程で栽培化が進んでおり、二つの起源地は別々であると結論付けている。 また、比較した3つの遺伝子の全てが、野生に近い優性遺伝子(コメの幅が細い、脱粒し易い、ねばねばが少ない)を持つイネが、東南アジア、特にインドネシアやフィリピンの熱帯ジャポニカに見られることを発見している。そして、ジャポニカの起源がインドネシアやフィリピンにあるとしている。
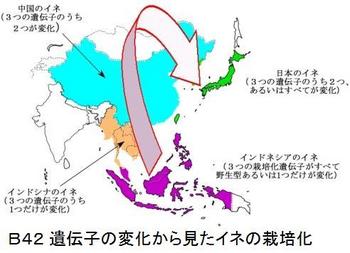 イネ栽培化の鍵である脱粒を抑制するタイプの遺伝子が中国と日本でしか見られず、東南アジアでは見つからなかったことから、次の結論を得ている。
イネ栽培化の鍵である脱粒を抑制するタイプの遺伝子が中国と日本でしか見られず、東南アジアでは見つからなかったことから、次の結論を得ている。
1)現在東南アジアで陸稲として栽培されている熱帯
ジャポニカがジャポニカの起源に近い。
2)熱帯ジャポニカが中国に伝わって長江流域で水田
化され、温帯ジャポニカが生れた。
3)温帯ジャポニカが更に日本に伝わった。
イネの起源地が中国でなく東南アジアの島々であったというのも衝撃的だが、熱帯ジャポニカから温帯ジャポニカが生れたというのも、熱帯ジャポニカの起源が明確でなかったから衝撃的な話だ。これら「遺伝子の変化から見たイネの栽培化」を図B42に、「3つの栽培化遺伝子の変化と分布」を図B43に示す(NIASのプレスリリース・論文(改変)より)。なお、栽培化遺伝子の変化は、qSW5 Wx qSH1、qsw5 Wx qSH1、qSW5 wx qSH1、qsw5 Wx qsh1、qsw5 wx qSH1、qsw5 wx qsh1の6種である。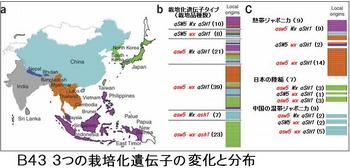
イネのDNA研究の先頭を走っているのが、農業生物資源研究所(NIAS)である。そのNIASが2008年7月、コメの大きさを決める遺伝子(qSW5)を発見したと発表した。それまでに見つけていたイネの脱粒性(穂からの籾のこぼれ易さ)の遺伝子(qSH1)と、炊いたコメのもちもち感を決める遺伝子(Waxy)と合わせて、三つの遺伝子の変化について、アジア15ヶ国のイネ、インディカ31種、ジャポニカ111種のDNA分析を行っている。
その結果、インディカでは、qSW5・qSH1・WaxyのDNAの変化はほとんど見られなかったが、ジャポニカでは、色々の変化のパターンが見られ、インディカとジャポニカは、別々の過程で栽培化が進んでおり、二つの起源地は別々であると結論付けている。 また、比較した3つの遺伝子の全てが、野生に近い優性遺伝子(コメの幅が細い、脱粒し易い、ねばねばが少ない)を持つイネが、東南アジア、特にインドネシアやフィリピンの熱帯ジャポニカに見られることを発見している。そして、ジャポニカの起源がインドネシアやフィリピンにあるとしている。
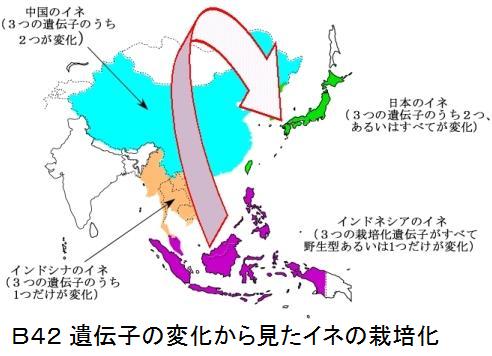 イネ栽培化の鍵である脱粒を抑制するタイプの遺伝子が中国と日本でしか見られず、東南アジアでは見つからなかったことから、次の結論を得ている。
イネ栽培化の鍵である脱粒を抑制するタイプの遺伝子が中国と日本でしか見られず、東南アジアでは見つからなかったことから、次の結論を得ている。1)現在東南アジアで陸稲として栽培されている熱帯
ジャポニカがジャポニカの起源に近い。
2)熱帯ジャポニカが中国に伝わって長江流域で水田
化され、温帯ジャポニカが生れた。
3)温帯ジャポニカが更に日本に伝わった。
イネの起源地が中国でなく東南アジアの島々であったというのも衝撃的だが、熱帯ジャポニカから温帯ジャポニカが生れたというのも、熱帯ジャポニカの起源が明確でなかったから衝撃的な話だ。これら「遺伝子の変化から見たイネの栽培化」を図B42に、「3つの栽培化遺伝子の変化と分布」を図B43に示す(NIASのプレスリリース・論文(改変)より)。なお、栽培化遺伝子の変化は、qSW5 Wx qSH1、qsw5 Wx qSH1、qSW5 wx qSH1、qsw5 Wx qsh1、qsw5 wx qSH1、qsw5 wx qsh1の6種である。

23-3.イネのDNAについて [23.稲の起源はスンダランド]
米国のノースカロライナ州立大学のM.Olsen氏は「Molecular Evidence on the Origin and Evolution of Glutinous Rice(モチ米の起源と進化についての分子レベルの証拠)」という論文を2002年に出している。アジアの105種の在来種のイネ、インディカ・熱帯ジャポニカ・温帯ジャポニカ、そしてモチ米・ウルチ米について、Waxy(粘性)遺伝子のタイプについて調査している。
この論文は最後に付表で、105種のイネのデータを全て載せている。私はこれらのデータをエクセルに取り込み色々層別してみた。すると、農業生物資源研究所のイネの起源と進化に対する結論より、より明確な答えを導き出すことが出来た。
これからの話を理解いただくため、ごく簡単にDNAの説明をしておく。DNAは色々な遺伝子が繋がっている紐と考えれば良い。一つの遺伝子は一つの仕事を持っている。例えば、米のWaxy遺伝子は、「米のねばねばを作る」という仕事であり、その仕事を為すための「指図書」と「設計図」を持っている。
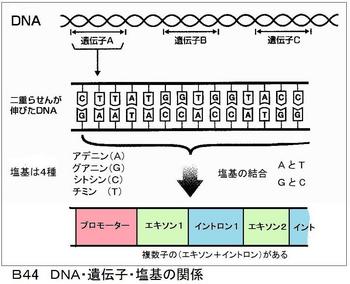 遺伝子の初めの部分は「プロモーター」と言われ、「いつ・何処に・何を」するかの「指図書」が書かれている。例えば「受粉後、胚乳に、ねばねばのデンプンを作る」等である。次にあるのが「エクソン」という部分で、作るでんぷんの「設計図」が書いてある。この設計図通りに、「リン酸―糖―塩基」のセットを組み立てると「アミロペクチンやアミロース」が出来る。リン酸と糖は全て同じだが、塩基には「A・G・C・T」の4種類があって、この4つの組合せの配列で異なった形質、たとえばアミロースの含有量の違ったものが出来る。
遺伝子の初めの部分は「プロモーター」と言われ、「いつ・何処に・何を」するかの「指図書」が書かれている。例えば「受粉後、胚乳に、ねばねばのデンプンを作る」等である。次にあるのが「エクソン」という部分で、作るでんぷんの「設計図」が書いてある。この設計図通りに、「リン酸―糖―塩基」のセットを組み立てると「アミロペクチンやアミロース」が出来る。リン酸と糖は全て同じだが、塩基には「A・G・C・T」の4種類があって、この4つの組合せの配列で異なった形質、たとえばアミロースの含有量の違ったものが出来る。
コンピーターが「0・1」の2つの組み合わせの配列で作動していることを考えると、4つの組合せで作られる生命体が、いかに複雑であるかが分かる。エクソンの後に「イントロン」という部分がある。この部分は「履歴書」みたいなもので、「米の粘りが弱い」などが書かれてある。図B44にそれらの概念図を示す。
M.Olsen氏の論文の内容は、英文で専門用語が多く、十分理解できていないが、大略すれば、「Waxy遺伝子の第一イントロンの先頭にある一つの塩基が、突然変異で変わると、胚乳のデンプンがねばりの体質に確実に変化する。そして、この突然変異が進化の起源の一つであり、それは東南アジアで起こった。ウルチ米からモチ米が発生した。」という内容である。
この論文は最後に付表で、105種のイネのデータを全て載せている。私はこれらのデータをエクセルに取り込み色々層別してみた。すると、農業生物資源研究所のイネの起源と進化に対する結論より、より明確な答えを導き出すことが出来た。
これからの話を理解いただくため、ごく簡単にDNAの説明をしておく。DNAは色々な遺伝子が繋がっている紐と考えれば良い。一つの遺伝子は一つの仕事を持っている。例えば、米のWaxy遺伝子は、「米のねばねばを作る」という仕事であり、その仕事を為すための「指図書」と「設計図」を持っている。
 遺伝子の初めの部分は「プロモーター」と言われ、「いつ・何処に・何を」するかの「指図書」が書かれている。例えば「受粉後、胚乳に、ねばねばのデンプンを作る」等である。次にあるのが「エクソン」という部分で、作るでんぷんの「設計図」が書いてある。この設計図通りに、「リン酸―糖―塩基」のセットを組み立てると「アミロペクチンやアミロース」が出来る。リン酸と糖は全て同じだが、塩基には「A・G・C・T」の4種類があって、この4つの組合せの配列で異なった形質、たとえばアミロースの含有量の違ったものが出来る。
遺伝子の初めの部分は「プロモーター」と言われ、「いつ・何処に・何を」するかの「指図書」が書かれている。例えば「受粉後、胚乳に、ねばねばのデンプンを作る」等である。次にあるのが「エクソン」という部分で、作るでんぷんの「設計図」が書いてある。この設計図通りに、「リン酸―糖―塩基」のセットを組み立てると「アミロペクチンやアミロース」が出来る。リン酸と糖は全て同じだが、塩基には「A・G・C・T」の4種類があって、この4つの組合せの配列で異なった形質、たとえばアミロースの含有量の違ったものが出来る。 コンピーターが「0・1」の2つの組み合わせの配列で作動していることを考えると、4つの組合せで作られる生命体が、いかに複雑であるかが分かる。エクソンの後に「イントロン」という部分がある。この部分は「履歴書」みたいなもので、「米の粘りが弱い」などが書かれてある。図B44にそれらの概念図を示す。
M.Olsen氏の論文の内容は、英文で専門用語が多く、十分理解できていないが、大略すれば、「Waxy遺伝子の第一イントロンの先頭にある一つの塩基が、突然変異で変わると、胚乳のデンプンがねばりの体質に確実に変化する。そして、この突然変異が進化の起源の一つであり、それは東南アジアで起こった。ウルチ米からモチ米が発生した。」という内容である。
23-4.イネの起源の解明に挑戦 [23.稲の起源はスンダランド]
 図B45は私が見出したイネの系統図で、Waxy遺伝子について、インディカ・熱帯ジャポニカ・温帯ジャポニカの3品種と、モチ・ウルチの2形質に分類している。英字はイントロンの先頭の塩基の種類を表わしている。カツコ付きの英文字は、Waxy遺伝子タイプを表わしている。
図B45は私が見出したイネの系統図で、Waxy遺伝子について、インディカ・熱帯ジャポニカ・温帯ジャポニカの3品種と、モチ・ウルチの2形質に分類している。英字はイントロンの先頭の塩基の種類を表わしている。カツコ付きの英文字は、Waxy遺伝子タイプを表わしている。  表B46に「Waxy(粘性)遺伝子の塩基配列」を示す。遺伝子タイプ「F」の塩基配列が一番上の列に書いてある。Fタイプと同じ塩基であれば「*」である。「-」は塩基がないことを示す。第1イントロンの先頭(1600番地、黄色)が、コメの粘性を大きく左右する塩基である。なお、コメの粘性に関わる塩基は、この塩基以外にWaxy遺伝子の第6エクソン等も関係しているが、第1イントロンの先頭が一番大きな効果があるそうだ。
表B46に「Waxy(粘性)遺伝子の塩基配列」を示す。遺伝子タイプ「F」の塩基配列が一番上の列に書いてある。Fタイプと同じ塩基であれば「*」である。「-」は塩基がないことを示す。第1イントロンの先頭(1600番地、黄色)が、コメの粘性を大きく左右する塩基である。なお、コメの粘性に関わる塩基は、この塩基以外にWaxy遺伝子の第6エクソン等も関係しているが、第1イントロンの先頭が一番大きな効果があるそうだ。 イネの起源は熱帯ジャポニカのウルチG(F)であると考える。これは、NIASの考え(qSW5 Wx qSH1)と同じである。熱帯ジャポニカがどのようして生れたかについては別途説明する。熱帯ジャポニカのウルチG(F)の1600番地の塩基が、突然変異でGからTに一文字変わっただけで、熱帯ジャポニカのモチT(G)が生れている。
熱帯ジャポニカのウルチG(F)が突然変異を起こし、温帯ジャポニカのウルチG(P)が生れる。この突然変異はWaxy遺伝子以外の遺伝子で起こっている。このG(P)を母親として、熱帯ジャポニカのモチT(G)を父親(花粉)として交配が起こり、温帯ジャポニカのモチT(G)が生れる。
この温帯ジャポニカのモチT(G)がウルチのT(G)に変化するのは、Waxy遺伝子の第1イントロンとは違う場所の突然変異であろう。粘性に一番関わりのある塩基がモチと同じだから、日本人が
 食する温帯ジャポニカのウルチ米は、モチ米ほどではないが、もちもち感がするのである。温帯ジャポニカのウルチT(G)の10
食する温帯ジャポニカのウルチ米は、モチ米ほどではないが、もちもち感がするのである。温帯ジャポニカのウルチT(G)の10番地の塩基が、突然変異でG→Aに変わり、ウルチ
T(M)が生れている。この品種の米がコシヒカリ
に代表される日本の米(qsw5 wx qsh1)であろう。
熱帯ジャポニカのウルチG(F)から、インディカのウルチG(F)が生れたと考える。世界のイネの研究者で、インディカの起源ついて明解な答えを見つけ学者はいない。インディカのウルチG(F)がどうして生れたかについては別途説明するとして、ここではインディカのウルチG(F)が生れた後を説明する。このG(F)を母親として、熱帯ジャポニカのモチT(G)を父親(花粉)として交配し、インディカのモチT(G)が生れる。
このインディカのモチT(G)が突然変異して、もしくは多種の野生イネと交配して生れたのが、インディカの変種T(R)である。インディカのT(R)は、モチもウルチもあり、両者の中間と言える。表B46の(R:ブルー)欄を見て頂くと、T(R)の多くの番地の塩基が変化しており、インディカの主なる品種(A,B)はこの塩基を持っている。インディカのモチ・ウルチT(R)とウルチG(F)の交配が起こり、インディカのウルチG(A)とG(B)が生れている。
23-5.イネの起源地を推定する [23.稲の起源はスンダランド]
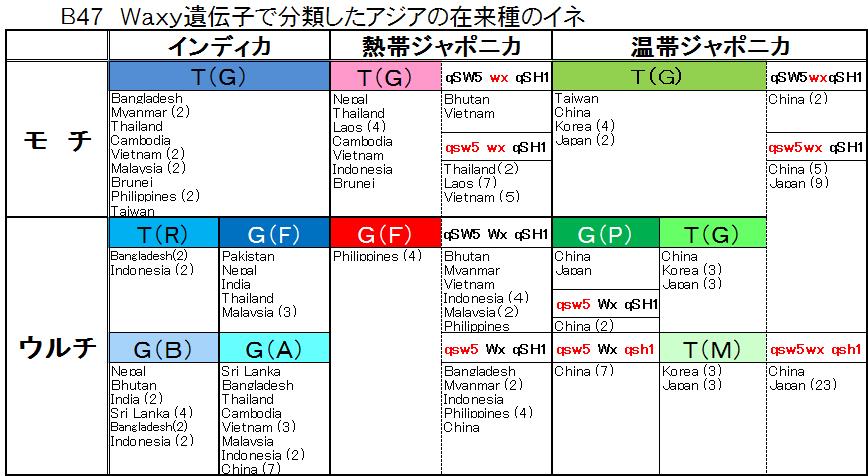 農業生物資源研究所(NIAS)のデータにはモチかウルチ化の区別はない。ただ、熱帯ジャポニカ(Tropical japonica)において、Waxy遺伝子が「wx」ならばモチ、「Wx」ならばウルチということが分かる。しかし、温帯ジャポニカ(Temperate japonica)は「Wx」ならばウルチとわかるが「wx」はウルチかモチか分からない。 私のイネの系統図(図B45)に、その在来種の国名をいれ、それにNIASの区別が明確な熱帯ジャポニカの「wx」と「Wx」、温帯ジャポニカの「Wx」のデータを付け加え、図B47を作成した。
農業生物資源研究所(NIAS)のデータにはモチかウルチ化の区別はない。ただ、熱帯ジャポニカ(Tropical japonica)において、Waxy遺伝子が「wx」ならばモチ、「Wx」ならばウルチということが分かる。しかし、温帯ジャポニカ(Temperate japonica)は「Wx」ならばウルチとわかるが「wx」はウルチかモチか分からない。 私のイネの系統図(図B45)に、その在来種の国名をいれ、それにNIASの区別が明確な熱帯ジャポニカの「wx」と「Wx」、温帯ジャポニカの「Wx」のデータを付け加え、図B47を作成した。ジャポニカの起源は、G(F)とqSW5 Wx qSH1の熱帯ジャポニカのウルチであり、起源地はフィリピン・マレーシア・インドネシア、ベトナムであると考えられる。 この考えに対し、佐藤氏は「イネの歴史」(2008年版)の中で、「この結論は、まず、考古学の成果とまったく合わない。東南アジア島嶼部における稲作の始まりは、考古学的にはせいぜい4000年ほど前のことである。長江流域における稲作の開始より数千年遅いのである。・・・これら島嶼部に現在すむ人々の先祖が伝わったのがせいぜい数千年前であるという人類学的な証拠もある。」と反対意見を述べている。
佐藤氏の反論は考古学者の発想だ。農学者・遺伝学者としての佐藤氏の考えはどうなのであろうか。論文を書いた正村純彦氏達(NIAS)の考え「三つの優性遺伝子は劣性遺伝子より古く、したがってこの三遺伝子を併せ持つ品種は、より古いタイプの品種ということになる」について、佐藤氏は「この推定は遺伝学的にも、また優性形質が劣性形質より古いタイプの形質である事実からみても妥当だと思われる」と述べている。イネの起源が東南アジア島嶼部にあったことは、遺伝学的に認められても、稲作の起源を考古学的には認められないのだろう。
23-6.スンダランドは存在した [23.稲の起源はスンダランド]
熱帯ジャポニカの起源地のフィリピン・マレーシア・インドネシア・ベトナムを地図で見ると、南シナ海の西部をぐるりと取り囲んでいる。現在中国が領有権を主張して、フィリピン・マレーシア・ベトナムと紛争している南沙群島の西側の海である。
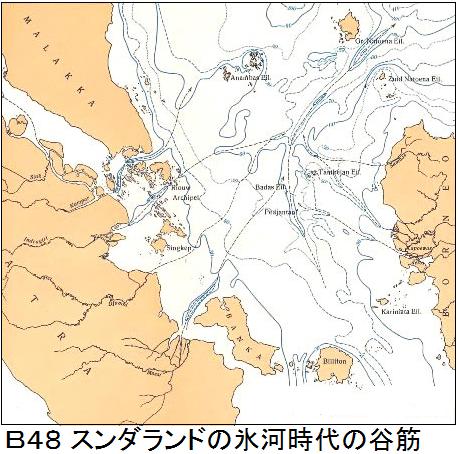
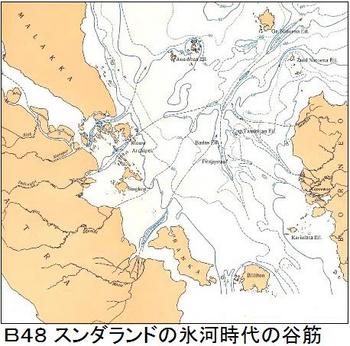 1920年インドネシアの海底地形の調査を行っていたオランダの地質学者のモーレングラーフ博士は、海底の等深線を作成した際に、海底に幾つもの谷筋を見つけた。それは、かつてそこに川が流れていたことを意味し、そこが陸地だったことを示していた。博士は今はなきこの幻の古代大陸を「スンダランド」と呼び、学会で発表して大きな反響を呼んだ。その後、海底のボーリング調査が行われ、花粉が見つかり、生態系が存在していたことが分かり、「スンダランド」が証明された。図B48にモーレングラーフ博士の作ったインドネシア周辺の海底の等深線図を示す。「Bibliography of Indonesia Geology, SUNDALAND」
1920年インドネシアの海底地形の調査を行っていたオランダの地質学者のモーレングラーフ博士は、海底の等深線を作成した際に、海底に幾つもの谷筋を見つけた。それは、かつてそこに川が流れていたことを意味し、そこが陸地だったことを示していた。博士は今はなきこの幻の古代大陸を「スンダランド」と呼び、学会で発表して大きな反響を呼んだ。その後、海底のボーリング調査が行われ、花粉が見つかり、生態系が存在していたことが分かり、「スンダランド」が証明された。図B48にモーレングラーフ博士の作ったインドネシア周辺の海底の等深線図を示す。「Bibliography of Indonesia Geology, SUNDALAND」
より引用。
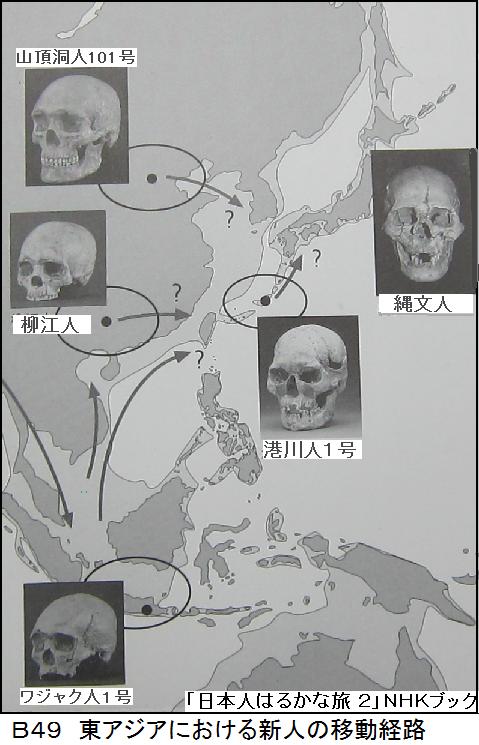
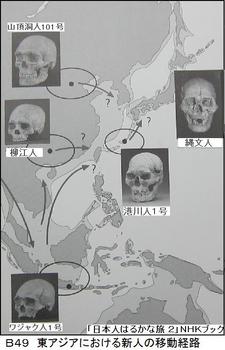 6万年前にアフリカを旅だった現代型のホモ・サピエンスは、4万年前にはスンダランドに到達している。そして3万年前には北東アジアに広がり、日本列島にも2万年前には到達している。もちろん、日本に到達したホモ・サピエンスは南回りだけでなく、北回りの人々もいた。図B49に、東アジアの新人の移動経路を示す。スンダランドは実在し、4万年前から現在の人類が住んだ所であった。
6万年前にアフリカを旅だった現代型のホモ・サピエンスは、4万年前にはスンダランドに到達している。そして3万年前には北東アジアに広がり、日本列島にも2万年前には到達している。もちろん、日本に到達したホモ・サピエンスは南回りだけでなく、北回りの人々もいた。図B49に、東アジアの新人の移動経路を示す。スンダランドは実在し、4万年前から現在の人類が住んだ所であった。
 1920年インドネシアの海底地形の調査を行っていたオランダの地質学者のモーレングラーフ博士は、海底の等深線を作成した際に、海底に幾つもの谷筋を見つけた。それは、かつてそこに川が流れていたことを意味し、そこが陸地だったことを示していた。博士は今はなきこの幻の古代大陸を「スンダランド」と呼び、学会で発表して大きな反響を呼んだ。その後、海底のボーリング調査が行われ、花粉が見つかり、生態系が存在していたことが分かり、「スンダランド」が証明された。図B48にモーレングラーフ博士の作ったインドネシア周辺の海底の等深線図を示す。「Bibliography of Indonesia Geology, SUNDALAND」
1920年インドネシアの海底地形の調査を行っていたオランダの地質学者のモーレングラーフ博士は、海底の等深線を作成した際に、海底に幾つもの谷筋を見つけた。それは、かつてそこに川が流れていたことを意味し、そこが陸地だったことを示していた。博士は今はなきこの幻の古代大陸を「スンダランド」と呼び、学会で発表して大きな反響を呼んだ。その後、海底のボーリング調査が行われ、花粉が見つかり、生態系が存在していたことが分かり、「スンダランド」が証明された。図B48にモーレングラーフ博士の作ったインドネシア周辺の海底の等深線図を示す。「Bibliography of Indonesia Geology, SUNDALAND」より引用。
 6万年前にアフリカを旅だった現代型のホモ・サピエンスは、4万年前にはスンダランドに到達している。そして3万年前には北東アジアに広がり、日本列島にも2万年前には到達している。もちろん、日本に到達したホモ・サピエンスは南回りだけでなく、北回りの人々もいた。図B49に、東アジアの新人の移動経路を示す。スンダランドは実在し、4万年前から現在の人類が住んだ所であった。
6万年前にアフリカを旅だった現代型のホモ・サピエンスは、4万年前にはスンダランドに到達している。そして3万年前には北東アジアに広がり、日本列島にも2万年前には到達している。もちろん、日本に到達したホモ・サピエンスは南回りだけでなく、北回りの人々もいた。図B49に、東アジアの新人の移動経路を示す。スンダランドは実在し、4万年前から現在の人類が住んだ所であった。
23-7.熱帯ジャポニカの起源はスンダランド [23.稲の起源はスンダランド]
 熱帯ジャポニカのウルチの起源地のスンダランドを図B50に示す。この地で生れた熱帯ジャポニカの子孫がフィリピン・マレーシア・インドネシア・ベトナムの地に残っていたことも納得出来る。 スンダランドは、モーレングラーフ博士の海底の等深線にあるように、現在のカリマンタン(ボルネオ)島やスマトラ島から川が流れ、そしてインドシナ半島のメコン川やチャオプラヤ(メナム)川も流れ込み、大デルタ地帯であったであろう。
熱帯ジャポニカのウルチの起源地のスンダランドを図B50に示す。この地で生れた熱帯ジャポニカの子孫がフィリピン・マレーシア・インドネシア・ベトナムの地に残っていたことも納得出来る。 スンダランドは、モーレングラーフ博士の海底の等深線にあるように、現在のカリマンタン(ボルネオ)島やスマトラ島から川が流れ、そしてインドシナ半島のメコン川やチャオプラヤ(メナム)川も流れ込み、大デルタ地帯であったであろう。 佐藤氏はベトナムのメコンデルタ地帯や、スマトラ島パレンバンのデルタ地帯で、水辺に群生する野生イネ、ルフィポゴンを観察している。その群生の大きさは百数十平方メートルにもおよび、その群生の島に上陸したと書いている。このような群生はカリマン島でも見られるという。そして、これらの野生イネは、まったくと言っていいほど、穂を付けず実をつけないそうである。
デルタの野生イネのルフィポゴンは、多年生で茎を伸ばしてその節から分枝をだして繁殖しており、種子で繁殖することはしていない。これを栄養繁殖というそうだ。佐藤氏の著書「イネの歴史」には、種子をつけないルフィポゴンの株をポットで育てた話があり、「予想だにしなかったことだが、その株は次のシーズンにはたくさんの穂をつけ、多量の種子をつけたのだった。私たちはこれに仰天したが、植物学の教科書には、植物たちは、乾燥、低温、養分不足などのストレスにあうと、とたんに花を咲かせて種子をつける「行動」にでると書かれてある」と述べている。
 2万年近く前、氷河期が終わった頃、スンダランドのデルタ地帯には、野生イネのルフィポゴンの先祖が群生を作っていたに違いない。その種子をつけない、ルフィポゴンの種子を人類はどうして手にいれたのであろうか。図B51に氷河期の終わりの海面上昇を示す。氷河期が終わった2万年前から現在までに海面は120メートル上昇している。この海面上昇により、スンダランドは7千年前に水没してしまうのである。
2万年近く前、氷河期が終わった頃、スンダランドのデルタ地帯には、野生イネのルフィポゴンの先祖が群生を作っていたに違いない。その種子をつけない、ルフィポゴンの種子を人類はどうして手にいれたのであろうか。図B51に氷河期の終わりの海面上昇を示す。氷河期が終わった2万年前から現在までに海面は120メートル上昇している。この海面上昇により、スンダランドは7千年前に水没してしまうのである。 デルタ地帯に群生していたルフィポゴンは、海面上昇による海水の侵入にストレスを、それ以上の身の危険を感じたであろう。そして、いっせいに穂を出し花を咲かせ、種子を実らせたと思われる。海岸線の後退は毎年少しずつ奥地へと広がり、ルフィポゴンの実りも毎年あったに違いない。こうして人類はイネの種子、コメを多量に手にいれたのである。
約1万年前には、ルフィポゴンの種子を栽培する人々も出て来たと考える。ただ、ルフィポゴンは水辺で栽培すると野生に戻り実をつけない。陸地で育てはなければ実をつけないのである。ルフィポゴンの子孫である熱帯ジャポニカが、多年草であるにも関わらず陸稲であるのは、野生にもっとも近かったためであろう。G(F)・qSW5 Wx qSH1の陸稲である熱帯ジャポニカは、スンダランドで生れたのである。熱帯地方は雨が多く陸稲として育てやすかったのであろう。



